En biología celular, los centríolos son una pareja de estructuras que forman parte del citoesqueleto semejantes a cilindros huecos. Los centríolos son orgánulos que intervienen en la reproducción celular, siendo una pareja de centríolos un diplosoma sólo presente en células animales. Los centríolos son dos estructuras cilíndricas que, rodeadas de un material proteico denso llamado material pericentriolar forman el centrosoma o COMT (centro organizador de microtúbulos) que permiten la polimerización de microtúbulos de dímeros de tubulina que forman parte del citoesqueleto. Los centríolos se posicionan perpendicularmente entre sí.
Cada centríolo está formado por nueve tripletes de microtúbulos formando un círculo. El más interno se llama microtúbulo A y está completo (compuesto de trece protofilamentos). A él se unen dos microtúbulos: el microtúbulo B que comparte tres protofilamentos con el A y el microtúbulo C, el más externo, que comparte tres protofilamentos con el B.
Los tripletes se unen entre sí gracias a una proteína llamada nexina, que conecta el microtúbulo A con el C del siguiente triplete. De cada triplete salen en forma de radios las fibrillas radiales, dejando una estructura denominada "rueda de carro" ó 9+0 por tener nueve tripletes externos y ninguno en el centro.
El proceso de formación ciliar en las células de diferenciación comprende la replicación del centríolo para originar múltiples procentríolos. Éstos crecen y migran hacia la superficie apical de la célula, en donde cada uno de ellos se convierte en un cuerpo basal. Desde cada uno de los nueve tripletes que forman el cuerpo basal crece un doblete de microtúbulos que produce una evaginación de la membrana apical. Esta proyección de la membrana contendrá los nueve dobletes periféricos que hay en un cilio maduro.
El material pericentriolar es un material denso y de naturaleza proteica que puede estar relacionado con la formación de microtúbulos. Esto es así ya que las células vegetales, que carecen de centríolos, también forman microtúbulos. Las células vegetales, en su lugar poseen una masa fibrosa difusa que tiene una composición similar al material pericentriolar.
El centríolo interviene en la división y movimiento cromosómico en la mitosis.

Cilios.-
Los cilios son expansiones celulares filiformes, de unos 0,25 µm de diámetro y unos 10 a 15 µm de longitud, que aparecen en las células animales y en algunos protozoos. Suelen disponerse densamente empaquetados, a modo de cesped, en las superficies libres de numerosas células, como las que forman los epitelios de los tractos respiratorios, de los conductos del aparato reproductor femenino de mamíferos o de las branquias de los peces y bivalvos. También aparecen en numerosos protozoos. Son estructuras que pueden moverse y su principal misión es la de desplazar fluidos, como ocurre con el mucus del tracto respiratorio, pero también empujan al óvulo a lo largo de las trompas de falopio hasta el útero o mueven el agua alrededor de las branquias. Los organismos unicelulares los usan para moverse ellos mismos o para arremolinar el líquido que les rodea y así atraer alimento. Una función del movimiento ciliar descubierta recientemente está implicada con el establecimiento de la lateralidad de determinadas estructuras de los vertebrados durante el desarrollo embrionario. El tipo de movimiento que realizan es de bateo, a modo de látigo, de manera sincronizada, produciendo una especie de ola que desplaza el fluido en una dirección paralela a la superficie de la célula.

Esquema que ilustra los modelos de movimiento propuestos para los cilios y los flagelos. En cada caso el flujo neto del fluido es diferente.
Se han observado numerosos cilios, denominados cilios primarios, que no funcionan como estructuras móviles. Prácticamente todos los tejidos animales estudiados, excepto las células sanguíneas, poseen cilios primarios: oviductos, neuronas, cartílado, ectodermo de las extremidades en desarrollo, células mesenquimáticas, ventrículos cerebrales, células epiteliales de los conductos urinarios, conductos prancreáticas, células hepáticas, e incluso células en cultivo. La mayoría de estos cilios no son móviles y se pensó que no eran funcionales. Sin embargo, se observó que la membrana ciliar tenía numerosos receptores y canales iónicos, por lo que se le asignó un papel sensorial. Por ejemplo, los receptores olfativos se encuentran en cilios dendríticos y los segmentos externos de los conos y bastones de la retina son en realidad cilios modificados. Algunos de los receptores están más densamente empaquetados en sus membranas que en el resto de la membrana plasmática de la célula. Además, existen numerosas moléculas en el interior del cilio primario que transducen estas señales. La mayor relación superficie/volumen hace que las respuestas intraciliares sean muy intensas frente a señales externas relativamente débiles. Además de sustancias químicas también pueden detectar movimientos de fluidos circundantes, actuando como mecanoreceptores.
Flagelos
Los flagelos son similares a los cilios pero mucho más largos, con unas 150 µm de longitud, y un poco más gruesos. Su principal misión es desplazar a la célula. Son mucho menos numerosos que los cilios en las células que los poseen. Su movimiento también es diferente puesto que no desplazan el líquido en una dirección paralela a la superficie de la célula sino en una dirección paralela al propio eje longitudinal del flagelo. Los flagelos son frecuentes en células móviles como ciertos organismos unicelulares y gametos masculinos.
Estructura
Los cilios y flagelos son estructuras complejas con más de 250 proteínas diferentes. Ambos contienen una estructura central de microtúbulos y otras proteínas asociadas, denominadas conjuntamente como axonema, rodeado todo ello por membrana celular. En su interior, además del axonema, se encuentran una gran cantidad de moléculas solubes que participan en cascadas de señalización y que forman la denominada matriz. Un axonema consta de 9 pares de microtúbulos exteriores que rodean a un par central. A esta disposición se la conoce como 9x2 + 2. El par central de microtúbulos contiene los 13 protofilamentos típicos, pero las parejas externas comparten protofilamentos. Los cilios primarios carecen de par central. A uno de los microtúbulos de cada par periférico se le denomina túbulo A y al otro túbulo B. El A es un microtúbulo completo mientras que el B contiene sólo 10 u 11 protofilamentos propios y 2 o 3 compartidos con el A.

Esquema donde se indican los principales componentes de la estructura de un cilio o un flagelo. En los cilios primarios el par central de microtúbulos está ausente.
Esta disposición se mantiene gracias a un entramado de conexiones proteicas internas. Al menos doce proteínas diferentes se han encontrado formando parte del axonema, las cuales están implicadas fundamentalmente en mantener la organización de los microtúbulos. Las parejas de microtúbulos externos están conectadas entre sí mediante una proteína denominada nexina. Los túbulos A de cada pareja están conectados por radios proteicos a un anillo central que encierra al par central de microtúbulos. En los microtúbulos externos aparece una proteína motora asociada llamada dineína que está implicada en el movimiento de cilios y flagelos.
Los microtúbulos se originan por polimerización a partir de una estructura localizada en el citoplasma celular periférico denominada cuerpo basal. La estructura del cuerpo basal es similar a la de los centriolos, es decir, 9 tripletes de microtúbulos que se disponen formando una estructura cilíndrica. Carece del par central (9x3 + 0). En cada triplete sólo uno de los microtúbulos contiene una forma completa y los otros dos comparten protofilamentos. Entre el cuerpo basal y el axonema del cilio existe una zona de transición que posee sólo los 9 dobletes típicos del cilio pero no el par central. Éste se formará a partir de una estructura llamada placa basal, localizada entre la zona de transición y el doblete interno. Los microtúbulos tienen sus extremos menos localizados en la punta distal de los cilios y flagelos. La parte del cuerpo basal más próxima al interior celular se ancla al citoesqueleto mediante estructuras proteicas denominadas radios ciliares
Además del axonema y sus proteínas asociadas se pueden encontrar otros tres compartimentos en los cilios, sobre todo en los cilios primarios. La membrana ciliar que, en los cilios primarios, contiene numerosos receptores y canales, consistente con la función sensorial. Otro compartimento es la matriz, la fase fluida que ocupa el interior ciliar. La matriz, además de ayudar a matener la estructura del flagelo, también tiene proteínas que transducen la señales generadas en la membrana. Otros dos compartimentos son la base y la parte más distal del cilio. En la base se encuentra el cuerpo basal y complejos proteicos desde los que parten y nuclean los microtúbulos del axonema. En la parte distal se encuentra un entramado proteico complejo donde aparecen proteínas asociadas a los microtúbulos que estabilizan los extremos menos.
Microfilamento
Los microfilamentos son finas fibras de proteínas de 3 a 7 nm de diámetro. Están compuestos predominantemente de una proteína contráctil llamada actina.
Los filamentos de actina o microfilamentos se sitúan en la periferia de la célula y se sintetiza desde puntos específicos de la membrana celular. Son los responsables de la forma y del desplazamiento celular. Están formados por proteínas globulares.
La asociación de los microfilamentos con la proteína miosina es la responsable por la contracción muscular. Los microfilamentos también pueden llevar a cabo movimientos celulares, incluyendo desplazamiento, contracción y citocinesis. en conjunción con los microtubulos le dan a la célula la estructura y el movimiento.
Organización
Los filamentos forman distintas proyecciones según la situación de la célula:
- Proyecciones dinámicas:
- a.- Lamelopodios (con forma de lámina) y filopodios (forma filamentosa y que censa el ambiente para decidir si la célula avanza no), que son estructuras que protruyen de la membrana celular y que permiten el movimiento de la célula.
- b.- Anillo contráctil: se forma cuando se está dividiendo la célula, una vez que los cromosomas se han separado, y estrangula la célula para dividirla en dos.
- Proyecciones estables: permanecen en el tiempo. Son por ejemplo, los paquetes de estereocilios (están en la superficie de las células pilosas del oído interno) u otros arreglos que permiten la contracción muscular.
Polimerización de Microfilamentos
La actina está situada en los bordes de la célula por lo tanto desde ahí se polimeriza. Comienza como respuesta a señales externas que le dicen a la célula la forma que tiene que adoptar. Lo primero que se forma es una especie de capuchón formado por proteínas especiales que son la ARP2 y la ARP3, junto con otras proteínas que fortalecen este capuchón y que forman el complejo ARP (proteína relacionada con actina). A partir del capuchón de unen los monómeros de actina para formar los protofilamentos. El lado menos tiene el capuchón por lo tanto el filamento crece solo en el sentido más. El capuchón puede unirse a otros filamentos para ramificarse.
Funciones de los microfilamentos
Tienen una misión esquelética. Son responsables de los movimientos del citosol. También son los responsables de la contracción de las células musculares. Por ejemplo, muchos tipos de células tienen microvellosidades, que son prolongaciones de la membrana plasmática que aumenta la superficie de contacto de la célula para mejorar el transporte de materiales a través de esta membrana. Las microvellosidades tienen haces de microfilamentos, los cuales se extienden y retraen como resultado del ensamble y desensamble de éstos.
Microtúbulos
Los microtúbulos son tubos cilíndricos de 20-25 nm de diámetro. Están compuestos de subunidades de la proteína tubulina , estas subunidades se llaman alfa y beta. Los microtúbulos actúan como un andamio para determinar la forma celular, y proveen un conjunto de “pistas” para que se muevan las organelas y vesículas. Los microtúbulos también forman las fibras del huso para separar los cromosomas durante la mitosis y la meiosis. Cuando se disponen en forma geométrica dentro de cilios y flagelos, son usados para la locomoción (autopropulsión) o para mover líquido circundante o partículas (motilidad).
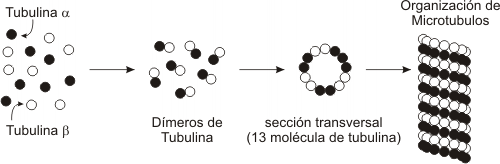
Fig. 6.4- Polimerización de la tubulina a partir de las tubulinas alfa y beta
Como mencionamos anteriormente la tubulina forma polímeros. La tubulina es una proteína globular, de la que existen dos polipéptidos distintos aunque similares, la alfa tubulina y la beta tubulina. La alfa y la beta tubulina se asocian y forman dímeros. En presencia de GTP, los dímeros de tubulina se unen y forman un tubo cuya parte central se mantiene vacía. Al igual que la actina F, los microtúbulos manifiestan polaridad, un extremo tiende a la polimerización o despolimerización a mayor velocidad (extremo +) y en el otro extremo ocurre lo mismo pero a menor velocidad (extremo).
Los microtúbulos se organizan a partir de centros organizadores especializados, que controlan su localización y orientación en cel citoplasma. El centro organizador principal en las células animales es el centrosoma, próximo al núcleo. El centrosoma esta formado por estructuras en forma de anillo que contiene otra tipo de tubulina, la gama tubulina. Estos anillos actuan como centros de nucleación (crecimiento) de microtúbulos. Los dímeros de tubulina se añaden al anillos de gama tubulina con una orientación específica, siempre el "extremo -" de cada microtúbulo queda dentro del centrosoma y el crecimiento se produce por el "extremo +" .

Extremos + y - de un microtúbulo
Las proteínas asociadas a los microtúbulos reciben el nombre de proteínas MAP (proteínas asociadas a los microtúbulos).
Por su localización, podemos clasificarlos en:
1. Citoplasmáticos (célula en interfase)
2. Mitóticos (fibra del huso)
3. Ciliares (en el eje de los cilios)
4. Centriolares (en cuerpos basales y centríolos)
Los microtúbulos citoplasmáticos son necesarios como vías de transporte de macromoléculas y organoides (vesículas, mitocondrias, etc.), intervienen dos proteínas motoras quinesina y dineína. En la neurona existe otra proteína motora asociada a los microtúbulos, la dinamina.
También establecen la forma celular. En las neuronas se hallan en las dendritas y en el axón, donde son esenciales para el crecimiento de éste último, que depende del alargamiento de sus microtúbulos. Este alargamiento es dependiente de la proteína motora dinamina, que provoca el deslizamiento de los microtúbulos, unos sobre otros.
En las neuronas se ha descubierto una MAP reguladora, denominada tau, que estabiliza los microtúbulos. En la enfermedad de Alzheimer, caracterizada por el deterioro neuronal progresivo, esta alterado el funcionamiento normal de esta proteína y por lo tanto se ve incrementada la inestabilidad de los microtúbulos imposibilitando el transporte axónico.
Los microtúbulos mitóticos movilizan los cromosomas durante la mitosis y la meiosis.
Los microtúbulos de cilias y flagelos crecen a partir de un cuerpo basal o cinetosoma de estructura idéntica a la de un centrosoma que actua como centro de nucleación de dímeros de alfa-beta tubulina. El cuerpo basal se encuentra por debajo de la membrana plasmática.
Existen diversas drogas que afectan a los microtúbulos, por ejemplo, la colchicina que se une a las tubulinas e impide su polimerización, lo que en definitiva produce la despolimerización de los microtúbulos. También pueden hacerse desaparecer los microtúbulos mitóticos mediante el uso de las drogas vinblastina y vincristina, que actúan de forma semejante a la colchicina, pero en forma selectiva, sobre los microtúbulos del huso mitótico. Por lo tanto estas drogas bloquean la división celular. Otra droga que produce los mismos efectos es el taxol, que impide la despolimerización de los microtúbulos, lo que induce su crecimiento descontrolado volviéndose imposible la división celular.
No hay comentarios:
Publicar un comentario